ДНК-МАРКЕРЫ
В СЕЛЕКЦИИ КЛЕВЕРА ЛУГОВОГО
Н. Н. Козлов, И. А. Клименко
Всероссийский научно-исследовательский институт
кормов имени
В. Р. Вильямса Российской академии
сельскохозяйственных наук, Россия
Во второй половине прошлого века широким фронтом
начались исследования молекулярных основ наследственности. Интенсивная работа в
этом направлении дала положительные результаты. В 50-х годах были разработаны
методы, позволяющие определять последовательность аминокислот в полипептидной
цепи белков и восстанавливать на этой основе нуклеотидную последовательность
транскрибируемой ДНК. А с появлением прямых методов ферментативного секвенирования
ДНК, предложенных М. Сингер в 1975–1977 гг. (Сингер, Берг, 1998), позволили
автоматизировать эту операцию. В 1980 г. был разработан метод для оценки
полиморфизма ДНК, получивший название RFLP (restriction fragments length polymorphism)
(Botstein, White, Skohnick, et al., 1980). С использованием этого метода было установлено,
что растения имеют 108–1010 нуклеотидов ДНК с огромным полиморфизмом,
который может быть положен в основу создания ДНК-маркеров и насыщенных
генетических карт. Несколько позже, в 1983 г., американскими биохимиками во
главе с K. B. Mullis (Mullis, Faloone, 1987) была открыта полимеразная цепная реакция (ПЦР)
— это техника быстрого и многократного увеличения небольших фрагментов ДНК. В
настоящее время на базе ПЦР создана плеяда методов, способных давать объективную
информацию о полиморфизме ДНК (Saha, Mian,
Zwonitzer et al., 2005). Цель этой процедуры —
получение картинки с амплифицированными продуктами ДНК объектов исследования (рис.
1).
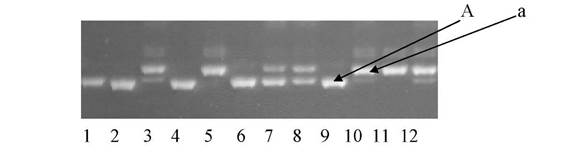
Рис. 1. Продукты
амплификации ДНК с использованием SSR праймеров, где 1,
2, 3 … n — выборка растений из
расщепляющейся популяции клевера лугового; А и а — полиморфизм одного из локусов
Последние достижения в области ДНК-технологий
послужили базой для кардинального изменения ситуации в теоретической и практической
селекции (Кочиева, 1999). Если в теоретическом плане маркирование генома стало
незаменимым инструментом при изучении функционирования генов, генетической
регуляции и фенотипической экспрессии, то их прикладное значение заключено в
маркировании селекционно-ценных признаков и свойств культурных растений с
широким использованием этой информации в маркер-вспомогательной селекции
(marker-assisted selection — MAS). Таким образом, впервые селекционер получил
инструмент, позволяющий вести селекцию на основе нуклеотидной
последовательности ДНК, то есть работать на основе полиморфизма
непосредственного носителя генетической информации.
Преимущество маркер-вспомогательной селекции (MAS), прежде всего, заключается в адекватности генетического
скрининга исходного материала, что приводит к повышению точности отбора, даже
если селективная аллель является рецессивной, а растения гетерозиготны. Это
приводит к сокращению количества циклов или даже этапов селекционной программы
и реальному ускорению селекционного процесса на несколько лет. А за счет более
раннего, хотя бы на год, начала использования сорта в производстве можно
получить такие дивиденды, которые существенно окупят все затраты на создание
данного сорта. По данным Pandey и Rajatasereekul (1999), более раннее (на 2
года) начало использования перспективного сорта риса может дать до 18 млн.
долларов прибыли. Нельзя недооценивать прямые и косвенные дивиденды, связанные
с возможностью проведения оценки отбора на ранних стадиях роста и развития
растений — вплоть до 5–7-дневных проростков, что дает возможность выбраковывать
значительное количество малоперспективного материала и экономить за счет этого финансовые
и материальные ресурсы.
Однако для широкого внедрения MAS
в селекционную практику необходим большой объем предварительных исследований,
связанных с маркированием селекционно-ценных признаков и свойств
культурных растений, а также с разработкой новых методов и схем селекционного
процесса.
Маркирование является наиболее сложным и трудоемким
подготовительным этапом MAS. При маркировании
необходимо выдержать ряд условий, которые предусматривают экономическую
обоснованность метода оценки ДНК-полиморфизма; тесную (≤ 1 сМ) связь ДНК-маркера
с селекционно-ценным признаком (Mohan, Nair, Bhagwat, at al. 1997); высокую воспроизводимость
техники скрининга ДНК-маркера в расщепляющихся популяциях.
Что касается экономической стороны способов скрининга
ДНК-маркеров, то уже сегодня их широкое разнообразие по стоимости и трудоемкости
позволяет выбрать вполне приемлемые методы, которые по цене сопоставимы с
массовыми методами определения содержания протеина, калия или других химических
элементов (Gu, Weeden, Yu, Wallace,
1995). В большей степени этим условиям удовлетворяют методы, основанные на ПЦР.
Более того, интенсивные поисковые работы в этом направлении привели к созданию
принципиально нового метода амплификации фрагментов ДНК. В этом случае вместо
традиционной полимеразной цепной реакции (ПЦР) в термоциклерах (приборы для постановки
ПЦР) используется метод геликаза-зависимой изотермической амплификации ДНК
(Helicase-dependent isothermal DNA amplification), требующей несколько
реактивов и инкубацию при 37 °C в течение одного–двух часов (Mariam Vincent, Yan Xu, Huimin Kong, 2004). Это
существенно упрощает и удешевляет скрининг маркеров в расщепляющихся популяциях.
Для успешной работы в области ДНК-маркирования необходима
насыщенная генетическая карта на основе ДНК-маркеров. Для целого ряда
культурных видов, в том числе и клевера лугового, такие карты уже созданы (Isobe et al., 2003, Sato et al., 2005, Herrmann et al., 2006).
Две из них принадлежат японско-российской творческой группе, включая
сотрудников ВНИИ кормов им. В. Р. Вильямса, а третья — австрийским
исследователям. При создании этих карт использовались различные методы оценки
ДНК-полиморфизма, основанные как на RFLP, так и PCR.
В современной синтетической селекции очень часто стоит
задача улучшения уже районированного сорта лишь по одному из признаков. В этом
случае используется донор и специальная схема беккроссирования, чтобы перенести
гены, ответственные за проявление селекционного признака и уменьшить долю ДНК-донора
у нового сорта. В данном случае MAS-селекция просто
незаменима, так как позволяет контролировать не только селекционный признак и даже
не только группу сцепления, в которой он находится, а полностью ДНК-донора и
реципиента. Конечно, это возможно лишь в случае, когда детально изучен
полиморфизм ДНК обоих родителей.







