|
|
МЕТОДЫ БИОТЕХНОЛОГИИ В
СОЗДАНИИ НОВЫХ СЕЛЕКЦИОННЫХ ИСТОЧНИКОВ КЛЕВЕРА ЛУГОВОГО (КЛЕВЕРА КРАСНОГО)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Число субкультур |
Число регенерантов |
Число выживших растений-регенерантов |
||
|
всего, шт. |
из них анеуплоидов, % |
шт. |
% |
|
|
111-У111 |
9136 |
0,9 |
134 |
98,5 |
|
1Х-ХУ |
5134 |
11,6 |
106 |
79,3 |
|
ХУ1-ХУ11 |
181 |
30,1 |
89 |
49,5 |
Анеуплоидные растения характеризовались пониженной фертильностью пыльцы, аномальным строением цветков (имели 1–4 пестика, 8–10 тычинок). Часть регенерантов была карликами и имела плохо развитую корневую систему.
Растения-регенеранты отличались от исходных растений по форме куста, длине, окраске и опушенности стеблей, наличию подковообразного пятна на листочках, размеру и количеству их в листе, форме, окраске и величине соцветий, строению и количеству головок в соцветии, продолжительности вегетационного периода.
Характер и количество измененных растений зависело от генотипа исходного растения и его происхождения. Наибольшей вариабельностью характеризовались клеточные культуры, полученные от растений гибридного происхождения (табл. 2).
2. Изменчивость признаков растений-регенерантов клевера лугового, полученных из клеточных линий различного происхождения
|
Клеточная линия |
Происхождение |
Проанализировано регенерантов |
Количество растений, % |
|||||
|
а |
б |
в |
г |
д |
е |
|||
|
Р-127 |
ВИК 7, 2n = 2х = 14 |
347 |
6,1 |
8,1 |
0,1 |
0,9 |
1,2 |
1,1 |
|
Тетра ВИК 7 |
В14 х Тетра ВИК 7, 2n = 4х = 28 |
401 |
14,8 |
1,2 |
34,1 |
14,5 |
49,5 |
0,1 |
|
ТР |
Тетра ВИК 7 х В-14, 2n = 4х = 28 |
308 |
26,3 |
0 |
26,8 |
19,6 |
59,4 |
0,1 |
|
Т-340 |
Тапио поли, 2n = 4х = 28 |
376 |
10,5 |
0 |
0 |
0,3 |
0 |
0 |
Примечание: а — нежизнеспособных, б — альбиносов, в — многолисточковых, г — многоголовчатых, в — с рассеченным цветком, е — с укороченной цветковой трубкой.
Таким образом, при первичном введении в культуру in vitro наблюдалась сомаклональная изменчивость. Однако при повторном введении в культуру in vitro измененных растений с признаками многолисточковость, рассеченность цветковой трубки, отсутствие пятна на листе были характерны для 100 % растений-регенерантов. Такие же результаты были получены при регенерации растений из эмбриогенной ткани, культивируемой на безгормональных средах.
Первое поколение (Р1) растений, полученное от переопыления исходных форм с растениями-регенерантами, не имеющими видимых изменений по изучаемым сомаклональным признакам, не отличалось от родительских форм. Однако среди растений второго (Р2) поколения наблюдалась большая вариабельность по этим признакам (рис. 1).
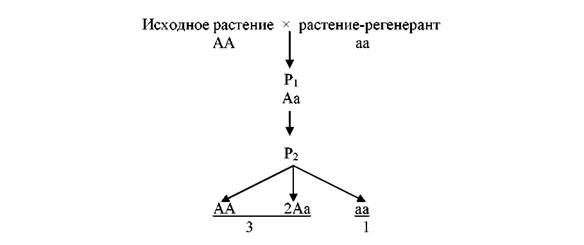
Рис. 1. Скрещивание исходного растения и растения-регенеранта клевера лугового (2n = 2х = 14)
Спектр изменчивости сомаклональных признаков растений Р2, полученных из семян одного соцветия, был идентичен спектру изменчивости растений Р0, регенерированных из клеток исходной клеточной линии. То есть возникшие в процессе культивирования сомаклональные изменения и не проявившиеся в Р1 поколении были рецессивными. Генетический анализ показал, что растения Р2, имеющие признаки альбиносов, многолисточковости и открытый тип цветения, составили примерно четвертую часть всех изученных в Р2 растений одной семьи. Совпадение фактически и теоретически ожидаемой численности растений по вышеназванным признакам с вероятностью 0,16–0,34 согласовалось с гипотезой, что в Р2 расщепление по фенотипам происходит в соотношении 3 : 1.
Одним из положительных признаков, наблюдаемых нами среди сомаклональных вариантов, является многолисточковость. Появление многолисточковых сомаклональных вариантов наблюдалось в большом количестве среди тетраплоидных регенерантов клеточных линий Тетра ВИК 7 и ТР (34,1 и 26,8 % соответственно, табл. 2). Значительно меньше их было идентифицировано среди растений клеточной линии Р-127 (4 из 347 растений).
Большой интерес для селекционеров могут представлять сомаклональные варианты клевера лугового, имеющие открытый тип цветка. Этот признак был генетически обусловлен и с вероятностью 0,34 наблюдался в 1/4 потомства растений-регенерантов от скрещивания, проведенного по вышеприведенной схеме. Цветок открытого типа имеет такой же длины цветковую трубочку (около 9 мм), что и исходное растение. Однако у него части венчика, образующие лодочку, не сросшиеся, что облегчает их опыление. О мутации цветка клевера лугового с несросшимися лепестками «дельта» сообщал Яблонски (1974), где эта мутация была выявлена среди 10 тысяч растений сорта Глория у единственного растения.
| | Оглавление Перейти на страницу |
стр. 91 |


Лаборатория молекулярно-генетических исследований кормовых культур
Лаборатория физиологии сельскохозяйственных растений
Взаимодействие:
Приморский НИИСХ





