|
|
Получение форм клевера лугового с повышенной устойчивостью к болезням методом генетической трансформации
Как отмечалось ранее, основой получения форм клевера лугового с повышенной устойчивостью к болезням методом клеточной селекции является высокая гетерогенность исходных суспензионных клеточных культур и, как следствие, большая сомаклональная изменчивость по ряду морфологических и физиологических признаков, как в положительную, так и в отрицательную сторону по сравнению с исходным генотипом. В связи с этим был разработан способ направленного создания методом генетической трансформации форм клевера лугового с одним из целевых признаков (устойчивость к болезням, вредителям и др.) и сохраняющих при этом все ценные селекционные признаки исходного образца. Для генетической трансформации использовали генотипы клевера лугового сортообразца Ранний 2 с высокой РС и показавшие высокую устойчивость к ионам алюминия при оценке in vitro, в вегетационном и полевом опытах. В опытах использовали штамм Agrobacterium tumefaciens LGV 3850 и штамм A. rhizogenes А4, содержащие векторные плазмиды pK22ac и pK22rs с маркерным геном npt11 (целевые гены ас-ар и rs-ap —гены синтеза дефензинов амаранта и редьки, повышающие устойчивость к корневым гнилям). Морфогенные культуры, инокулированные агробактериями, помещали на агаризованную среду Гамборга В5 с 2 мг/л БАП и добавлением 50 мг/л канамицина (селективного фактора для отбора клеточных культур клевера лугового с встроенными генами npt11) и 500 мг/л цефотаксима для подавления роста агробактерий. Канамицин добавляли в среды в продолжение всего процесса трансформации и поддержания in vitro коллекции трансформированных морфогенных культур клевера лугового. Субкультивирование на средах с цефотаксимом проводили каждые 3–4 недели, снижая постепенно его концентрацию до 0 мг/л до полной элиминации агробактерий. Подтверждение наличия генов nptII (маркерный), ac-ap (целевой) в полученных растениях определяли методом ПЦР. При этом очень важным моментом является то, что анализ проводили у растений, выращиваемых на среде Гамборга без добавления клафорана (цефотаксима) при условии отсутствия агробактериальной инфекции, что исключало из анализа нетрансгенные растения, дающие положительный результат по ПЦР за счет остаточной контаминации агробактериями. В связи с тем, что в исследуемых растениях могут присутствовать собственные гены пептидных антибиотиков, гомологичные введенным трансгенам, для идентификации трансгенных растений методом ПЦР мы использовали праймеры, комплементарные промоторной области 35S вируса мозаики цветной капусты и концевому участку гена ac-ap. Были проанализированы растения из следующих вариантов опыта:
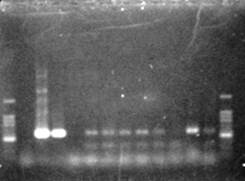
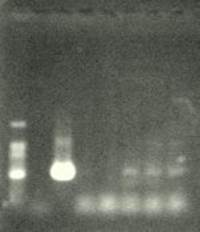
ПЦР-анализ с праймером npt1/npt2 при температуре отжига 50 ºС показал амплификацию всех тестируемых образцов, кроме № 7, с единственным специфичным фрагментом на уровне свечения плазмиды размером 580—590 пар нуклеотидов (рис. 2). В контрольном образце исходного растения (дорожка № 5 на рис. 2) амплификацию не наблюдали. При ПЦР-анализе исследуемых образцов на наличие целевого гена дефензина амаранта (ac) установлено, что при использовании комбинации праймеров 35S/ac при температуре отжига 52 ºС наблюдали амплификацию образцов, трансформированных штаммом агробактерии с геном дефензина амаранта при отсутствии амплификации в контрольном нетрансгенном растении (рис. 3 — дорожки 6,7,8, образцы LGVac2; LGVac15П; A4acI 200), что свидетельствует о наличие встраиваемых генов в геноме изученных образцов
1 2 3 4 5 6 7 8 9 10 11 12 13 14
Рис. 2. Результаты ПЦР-анализа растений клевера лугового с праймерами npt1/npt2 и плазмидой pK22rs (t отжига 50 ºС) лунки: 1 и 14 — маркер молекулярной массы; 2 — вода, 3 — плазмида pK22rs, 4 — плазмида pK22rs + ДНК исходного образца (контроль — Р8), 5 – контроль Р8; 6–13 — ДНК растений-трансформантов (№ 1, 2, 3, 4, 6, 7, 8, 9).
1 2 3 4 5 6 7 8 Рис. 3. Результаты
ПЦР-анализа с праймерами 35S/ac растений клевера лугового,
трансформированных штаммом, содержащим вектор с геном ac лунки: 1 — маркер
молекулярной массы, 2 — вода, 3 — плазмида pK22ac, 4 — pK22ac +
ДНК исходного образца (контроль — Р8), 5 — ДНК исходного образца (Р8), 6–8 (№
1, 2, 6) Все трансформированные морфогенные культуры клевера лугового сохраняли признак кислотоустойчивости на агаризованной питательной среде Гамборга В5 с 50 мг/л ионов алюминия после длительного (в течение более чем 56 пассажей) культивирования на среде с селективным фактором канамицином. Результаты представлены в таблице 6. Изученные клоны четырех генотипов клевера лугового сохранили признак устойчивости к А13+ на уровне исходных форм, а клоны трех генотипов A4rs21 I, LGVac15П, LGVac2 даже существенно превышали их по массе образовавшихся корней 72,1, 54,6 и 125 % соответственно. При этом длина корней была больше лишь у двух генотипов (LGVac15П и LGVac2). Существенное превышение массы морфогенной ткани отме чалось у клонов A4rs21 I (на 93,4 %). Средняя масса побегов трансгенных клонов превышала таковую исходных образцов у трех генотипов A4rsК7 11-2, A4rs21I и LGVac15П на 56,9; 38,5 и 32,2 % соответственно.
6. Кислотоустойчивость трансгенных морфогенных культур клевера лугового
Таким образом, полученные данные, и в том числе по способности к ризогенезу изученных клонов на селективной среде, свидетельствуют о сохранении признака кислотоустойчивости растениями-регенерантами в процессе генетической трансформации. Разработанный способ генетической трансформации растений селекционно-ценных образцов клевера лугового позволил получить растения-регенеранты со встроенными генами — маркерным (канамицинустойчивость) и целевым (ген синтеза дефензина амаранта, повышающего устойчивость растений к корневым гнилям). Использование для ПЦР-анализа ДНК, выделенной из листовой ткани асептических растений-регенерантов, образовавших корни на среде с 50 мг/л канамицина, позволило выявить трансгенные растения на более ранних этапах культивирования до высадки в почву. Кроме того, использование при генетической трансформации способа прямой регенерации растений, исключающего возникновение сомаклональной изменчивости, позволило: 1) сохранить селекционно-ценные признаки исходного образца (кислотоустойчивость), 2) поддерживать в культуре in vitro более 50 пассажей трансгенные морфогенные культуры с высокой регенерационной способностью, 3) получить неограниченное количество трансгенных растений-регенерантов.
|
Лаборатория молекулярно-генетических исследований кормовых культур Лаборатория физиологии сельскохозяйственных растений Взаимодействие: Приморский НИИСХ
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||